第4話「塩化セシウム平衡密度勾配遠心法」
籔井教授 さて、今日はウイルス精製の最終段階である塩化セシウム平衡密度勾配遠心法の話をしよう。超遠心によるウイルス精製の流れで行くと、③の段階になります。
超遠心によるウイルス精製の流れ
① 分画遠心によるペレッティング
↓
② ショ糖密度勾配速度ゾーン遠心法
↓
③ 塩化セシウム平衡密度勾配遠心法
早斐 なるほど、さらに③の段階があるのと、②の段階で終了というのとでは違うのですね。ところで、精製を②の段階で終了させて、最終精製ウイルスとすることもありますか?
籔井教授 ありますよ。不活化ワクチンのためにウイルス粒子を精製するのであれば、ショ糖密度勾配遠心法でウイルス粒子の精製を完了させるでしょう。塩化セシウム平衡密度勾配遠心法を使うと、目的によっては必要以上にウイルス粒子が細かく分画されてしまうことがあります。
早斐 前回の講義(第3話)を思い出したわ。ロタウイルス粒子は、カプシドが3層構造をとる完全粒子(triple-layered particle: TLP)と2層構造をとる2層粒子(double-layered particle: DLP)がありましたね。
籔井教授 そうです。ロタウイルスの不活化ワクチンを作ろうと思えば、二つの粒子が混ざっていてもかまわないでしょう。生ワクチンは接種された人の腸管内で増えて十分な抗原量となるので、培養液1 mLから一人分のワクチンが作られれば実用化が可能です。一方、不活化ワクチンとなると、十分な免疫応答が得られる抗原量をあらかじめ接種する必要があります。しかも不純物をできるかぎり少なくしなければならない。つまり、ウイルスの精製が必要なわけです。その一方で、実用化の見地からすると一人分のワクチンを作るのにどれくらいのコストがかかるかという関門があります。
早斐 なるほど。最終的に培養液何mLから一人分のワクチンが作られるか、ということが肝心なわけですね。精製過程から逆算して、1 Lから一人分のワクチンしか作れないとなると、確かにこれはちょっと厳しいかなという気がします。
籔井教授 さて、平衡密度勾配遠心法 (equilibrium density gradient centrifugation) は、ショ糖密度勾配遠心法とは原理が異なります。ショ糖密度勾配遠心法でカギとなるS値は関係しません。平衡密度勾配遠心法では、ある固有の浮上密度 (buoyant density) をもつウイルス粒子が遠心中に沈降していく途上で、溶媒の密度に一致したところでそれ以上沈降も浮上もしなくなります。つまり平衡状態に達するわけです。したがって、遠心を続けていくと、最初に溶液中のどこにウイルス粒子があってもこの等密度の場所に集まってきてバンドを形成します。このことを利用して、ウイルス粒子の精製を行います。
伊藤 ここに、沈降速度を出す計算式があります。これは沈降速度、つまり遠心チューブ中の粒子が遠心力の働く方向に一秒間あたり何センチメートル進んで行くかを示す計算式です。

v:沈降速度(cm/s)、d:粒子の直径(cm)、ρp:粒子密度(g/ml)、ρl:溶媒密度(g/ml)、
η:溶液の粘度(cm・s)、r:回転半径(cm)、ω:回転角速度(rad/s)
この式で、粒子の密度(ρp)と溶媒の密度(ρl)が等しくなると、(ρp-ρl)の項の値が0となります。その結果、沈降速度V=0となるので、沈降が止まることになりますね。
籔井教授 理屈はこのくらいにして、具体例を紹介しましょう。具体例は、今ちょうど早斐さんが思い出してくれたロタウイルスの2つの粒子、完全粒子TLPと2層粒子DLPを分離する話です。この2つは塩化セシウム溶液中での浮上密度が違います。TLPが1.36 g/mL、DLPが1.38 g/mLです。
早斐 完全粒子の方が軽いのですね。
籔井教授 ウイルスのゲノムである二本鎖のRNAの密度はだいたい1.7 g/mL、単純タンパク質の密度は1.3 g/mLですから、TLPにもDLPにも粒子1個当たり同じ量の核酸が入っているとすると、タンパク質からなるカプシドの層が1つ多いTLPの方の密度が少しだけ小さくなります。
二種類のロタウイルス粒子
| 表面に凹凸がある | 表面が平滑である | |
 |
 |
|
| DLP 2層粒子 | TLP 完全粒子 |
| 粒子 | 分子量 (MDa) |
核酸含量 (%(w/w)) |
タンパク質含量 (%(w/w)) |
浮上密度 (g/mL) |
| DLP 2層粒子 | 約 60 | 19 | 81 | 1.38 |
| TLP 完全粒子 | 約 100 | 11 | 89 | 1.36 |
伊藤 ロタウイルス精製の具体例についてお聞きしたいのですが、塩化セシウム平衡密度勾配遠心にかける前の出発材料は、1つ前のステップであるショ糖密度勾配遠心をスウィングロータ SW 41 Tiを使って行ったときの最終産物、つまり第三話でのBand Aのフラクションですね。このフラクションをどうやって回収するかというお話をうかがえませんか?
籔井教授 確かに、前回はスウィングロータ SW 41 Ti で38,000 rpm、4時間遠心した後、0.5 mLずつ21本のフラクションにして、採取した後の各フラクションをアッセイした結果を図で示しただけでしたね。早斐さんなら、Band Aのフラクションとして、どこからどこまで取ってプールしますか?
早斐 フラクション5から7にします。
籔井教授 なかなか思い切りがいいですね。
早斐 だって、次の平衡密度勾配遠心のステップがあるでしょ。フラクション5から7の3フラクションをプールして1.5 mL、これが6本あるので、最終的に9 mLになってしまいます。よくばって、フラクション4から8をプールすると、最終的には15 mLの出発材料になってしまいますものね。
籔井教授 結構。ところで、そのままで次のステップにもっていけますか?
早斐 あっ、スクロースを除かなければいけません。Band Aのフラクションには55%くらいのスクロースが入っていますから。
籔井教授 どうやって、除きますか?
早斐 一晩PBS(Phosphate Buffered Saline: リン酸緩衝生理的食塩水)に透析するか、少しうすめて超遠心でペレットにするかします。
籔井教授 次のステップである平衡密度勾配遠心には最低でも一晩かかりますから、超遠心でのペレッティングが速くていいですね。前にもお話しましたが、ペレッティングによる濃縮はウイルス粒子を扱うときの基本操作です。この操作で緩衝液中のスクロースを除くとともに、濃縮もできるわけです。
ウイルスのペレッティングによる濃縮はウイルス粒子を扱うときの基本操作です。
ところで、ペレッティングの前にどれくらいうすめますか?
早斐 スクロースの濃度を少なくとも20%より小さくすれば、ロタウイルス粒子も通常の遠心条件とあまりかわらずにペレッティングできると思います。そこでスクロースの濃度を20%未満にしたいのですが、そのためには少なくとも3倍に希釈する必要があります。この先は、次にどんなロータを使うかによります。私なら、ショ糖密度勾配遠心に使ったスウィングロータ SW 41 Tiを使います。

| 最高回転数 (rpm) |
最大遠心力 (xg) |
k-ファクタ | チューブサイズ (mm) |
公称容量 (本数 × mL) |
| 41,000 | 288,000 | 124 | 14×89 | 6×13.2 |
ウイルス液が9 mLですから、スクロースの濃度を20%未満にするためには、3倍の27 mLにすればいいわけですが、SW 41 Ti用のポリプロピレンチューブの容量は最大12 mL(公称容量は13.2 mL)ですので、2本では少し足りず、3本使うことになります。結論として、先ほどの9 mLのウイルス液に27 mLのPBSを 50 mLファルコン遠心管の中でよく混和して36 mLとしてから、SW 41 Ti用の3本のポリプロピレンチューブに12 mLずつ分注します。
伊藤 ピペットで正確に分注して、目視でチューブの水位が同一であれば、天秤を使ってバランスをとる必要はありません。遠心チューブは、1、3、5あるいは2、4、6の位置にあるバケットに入れていただきますが、残りの3本のバケットは空のままであっても必ず6本ともロータにかけてください。

早斐 はい、わかりました。ところで、ペレッティングの遠心条件は、スウィングロータ SW 41 Tiで38,000 rpm、1時間でいいでしょうか?
籔井教授 いいですよ。第一の目的はウイルス粒子をペレットにすることですから、SW 41 Tiであれば38,000 rpmで1時間の遠心でいいでしょう。前にも同様の話をしましたが、遠心力を小さくすると、ポリプロピレン製の遠心チューブの寿命が長くなります。たとえば、36,000 rpmで1.5時間にしただけでも、ポリプロピレン製の遠心チューブにクラックが入る頻度が大きく減ります。
ところで、ペレットは最終的にチューブあたり1 mLになるようにPBSに懸濁してください。ウイルスのペレッティングによる濃縮にあたって、注意すべきポイントがありましたね。思い出してください。
1. 濃縮しすぎに注意!目安はペレットを遠心前の容量の10分の一くらいの溶液に懸濁することです。
2. ペレットを懸濁するときは、あせらず時間をかけ、しっかりと。
要は、できるだけ細かく均一な懸濁液を作ること。
第一のポイントについてですが、スクロース濃度を下げるために希釈したので容量は36 mLになっています。しかし、希釈する前でも 9 mLのウイルス液でしたので、最終的に3 mL(チューブあたり1 mL)に懸濁するので、このポイントはクリアしています。出発材料がショ糖密度勾配遠心のフラクションなので、本来大きな塊(かたまり)はないはずですが、ペレットにすると凝集塊がでてくることがあります。ですから超遠心後には、第二のポイントである、できるだけ細かく均一な懸濁液を作ることにも注意を払ってください。
籔井教授 さて、いよいよ最後の塩化セシウム平衡密度勾配遠心です。ここでは、スウィングロータ SW 55 Tiを使ったときの遠心条件を示しましょう。

| 最高回転数 (rpm) |
最大遠心力 (xg) |
k-ファクタ | チューブサイズ (mm)*1 |
公称容量 (本数 × mL) |
| 55,000 | 368,000 | 148 | 13×51 | 6×5 |
塩化セシウム平衡密度勾配遠心法
- ウイルス粗精製溶液を調整する。
- スウィングロータ SW 55 Ti用のUCチューブ(公称容量5 mL)の底に55%(w/v)塩化セシウム溶液、次に、40%(w/v) 塩化セシウム溶液を重層する。
- 1で用意したウイルス粗精製溶液をチューブ1本当たり、0.5~0.8 mLを重層する。
- チューブをロードしたバケットをSW 55 Tiに載せ、 46,000 rpm (257,000 xg*)で16~20時間遠心する。
- 遠心後、チューブの底から3分の一付近に形成された青白い色をした近接する2本のバンドが精製ウイルス粒子である。上方のバンドがTLP、下方のバンドがDLPである。
- 上方のTLPのバンド直前まで、上から先端にパスツールピペットをつけ液を静かに吸引していく。それぞれのバンドは目視によりツベルクリン用の1 mLの注射器で回収する。
- 回収したそれぞれのバンド(0.5 mL程度におさまるはず)をPBSで8倍に希釈し(およそ4.2 mL程度におさまる)、新しい、ポリプロピレン製のSW 55 Ti用遠心チューブ(PPチューブ)に移す。
- チューブをロードしたバケットをSW 55 Tiロータに載せ、 40,000 rpm (194,000 xg)で1時間遠心する。上清を静かに吸引し、ペレットを1本あたり0.5-1.0 mLのPBSに懸濁する。これが最終精製標品になる。

* ここには最大遠心力を示していますが、ベックマンのウェブサイトに遠心力の計算機があり、ロータと回転数を指定すると、すぐに平均および最大遠心力を返してくれる。
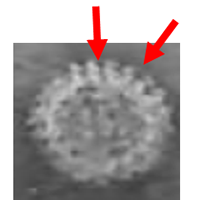
伊藤 塩化セシウムでの平衡密度勾配遠心後のチューブに見られる白っぽいモヤモヤした浮遊物はウイルスとは関係ない不純物だと思うのですが、その直下のウイルスバンドと同じように青白く見えるバンドは何なのでしょうか。
籔井教授 いいところに目をつけましたね。さらに言うと、TLPやDLPのバンドよりもさらに太い、つまり量としてたくさんあるところが気になるところなのです。密度もずっと軽く、核酸の存在を示す260 nmでの吸収も強くないので、単純タンパク質が主体だろうと推定されます。実は、この実験例ではないのですが、同様の塩化セシウムでの平衡密度勾配遠心後に、この部分を回収して電子顕微鏡で観察したものがありますので、その写真をお見せしましょう。

伊藤 ほとんどのウイルス粒子は中が黒くなっている中空のウイルス粒子ですね。
籔井教授 そうです。粒子の中に核酸がないので、染色液が入り込んだ結果、黒く染色されています。
早斐 中央のあたりにTLPも1個、そのちょっと下には中空ウイルス粒子に接する位置に少しいびつになったDLPも見えますね。
籔井教授 TLPやDLPは密度が高いので、本来はもっと下の方のバンドになるのですが、現実には、このように中空粒子と一緒に巻き込まれて存在することもまれではありません。絵で描くと簡単ですが、実験して論文の中で証拠として示そうとすると大変なのですよ。ロタウイルスの名付け親である英国のフルーエット(Thomas H. Flewett: 1922-2006)の弟子の一人にエリアスという人がいましたが、この人がTLPとDLPの二つのタイプの粒子を塩化セシウムでの平衡密度勾配遠心で分画する実験をやっています。
実に500個以上の粒子を観察して、密度1.36 g/mL のフラクションにはDLP 1個に対してTLPが69個存在すること。一方、密度が1.38 g/mLのフラクションにはDLP 128個に対してTLPが1個存在することから、TLPの密度が1.36 g/mLであり、DLPの密度が1.38 g/mLであることを示しました(Elias, M.M., Separation and infectivity of two particle types of human rotavirus. J Gen Virol 37: 191-194, 1977)。
早斐 なんだか回りくどいわね。でも教授の電子顕微鏡写真に写っているロタウイルス粒子数は30もありませんから、500個を調べるというのは、結構、大変そうですね。
籔井教授 エリアスさんは、早斐さんがめんどうでやりたくないと言っていた、これらの粒子の存在するフラクションの感染性を実験しています。その結果、TLPのフラクションにはDLPのフラクションの1000倍以上の感染性があることからTLPが感染性粒子であることを結論付けたいようでした。
早斐 わずかに存在するDLPのフラクションの感染性が、DLPにもわずかな感染性があるためなのか、わずかに混在しているTLP粒子によるものなのか決定する証拠がなかったわけですね。でも不完全な粒子であるDLPには感染性はありませんよね。
籔井教授 そのように理解されています。ただ、その後、誰かが証明したということでもありません。TLP粒子には、1977年当時は知られていなかったVP4からなる表面のスパイクタンパク質があり、これが宿主細胞のウイルスレセプターに結合して感染が開始されると理解されるようになりました。そこで、宿主細胞への感染を開始するのに必要なVP4もVP7もないDLPに感染力があるかどうかを、もはや問題にする人がいなくなったわけです。
早斐 そんな風に言われると、実験的証拠は変わらず、概念や考え方が変わっただけのような気がしますけど、そんなことでいいのですか?
籔井教授 そんなことでいいのかと早斐さんから突っ込まれましたので、科学的研究とは何かと言うおおもとになるところを考えてみましょう。第一は、研究と言うのはその時代にとって意味があり、かつ、その時代の技術で解くことができる問題を扱うということ。第二は、何を持って解けたとするかは、その分野のエキスパートが納得すれば、しかも仮に全員でなくても大多数が納得すれば、「解けた」あるいは「分かった」ということになるということです。これが自然科学研究の原則です。極論すれば、すべては仮説であり、そうではないと反証されるまで正しいということになります。
伊藤 科学哲学的論議が高じていかないように、最後に現実的な質問をさせてください。最近はウイルス粒子の浮上密度のことをあまり問題にしなくなったような気がしますが、どうしてなのですか?
籔井教授 話を具体的にするためと私の専門領域ということで、胃腸炎ウイルスを例にしてみましょう。ゲノムの核酸を直接検出する核酸診断法がなかった時代には、電子顕微鏡でウイルス粒子を検出するparticle virologyというのが主流でした。ところが、ロタウイルスのように形態的に診断の助けとなるような明確な特徴があればいいのですが、多くの胃腸炎ウイルスは、電子顕微鏡ではただの小型の球状粒子に見えます。細胞内でも、糞便内でもいろいろなものが電子顕微鏡で見ると小型の球状粒子なのです。そこで、これらの粒子が内部に核酸を持ったウイルス粒子であるとすると、単純タンパク質よりも重い一定の浮上密度を持っていることに着目して、同定する手段にしたのです。
伊藤 最後に、もう1つだけ。浮上密度はウイルス粒子の核酸含量で決まるのですか?
籔井教授 核酸含量は大きな要素です。ただそれだけではないようです。たとえば、ピコルナ科ウイルスでよく研究されているのですが、浮上密度にはウイルスRNAとセシウムイオンとの反応や水和の程度が影響することが知られています。(Rowlands DJ, Sangar DV, Brown F. Buoyant density of picornaviruses in caesium salts. J. Gen. Virol. 13):141-152, 1971)
関連ページ
|
|
超遠心機 ベックマン・コールターは1947年に初めて超遠心機(超遠心分離機)を販売開始し、現在では高い遠心力・回転数はもちろん、バイオセーフティモデルやリモートコントロール機能などを備えた超遠心機を取り揃えております。様々なチューブや容量に対応するロータやアクセサリ類も取り揃え、ウイルスの高純度精製やエクソソーム分離、その他幅広いアプリケーションのご要望にお応えします。 |

